
DRAFTING…
| Extracts From Christoph Parker and Hazel A. Oxenford, 2014. Environment and development in coastal regions and in small islands, Coastal region and small island papers 3: Barbados. 6pp. Available earlier at http://www.unesco.org/csi/pub/papers/parker.htm* *That URL is not currently functional. The same info., without colour in the figures and a few details about the monitoring sites can be found in Christoph Parker and Hazel A. Oxenford, 1998. Barbados.pp 177-186 in UNESCO, 1998. CARICOMP – Caribbean coral reef, seagrass and mangrove sites. Coastal region and small island papers 3, UNESCO, Paris, xiv + 347 pp. SUMMARY: The CARICOMP sites on Barbados include a reef site near Holetown on the west coast and, 25 km away on the south coast, a seagrass site (St. Lawrence Lagoon) and a mangrove site (Graeme Hall Swamp). The 32 ha Graeme Hall Swamp is fringed by red and white mangroves and is greatly impacted by a large population and several channel-dredging projects. The seagrass site is located in a shallow lagoon that is protected from high energy waves by a reef rubble bank; turtle and manatee grasses dominate. Shallow fringing reefs are fairly extensive along the west coast of Barbados. The reef site, the Bellairs fringing reef, extends 300 m seaward from the shoreline and is very shallow, with the base of the reef at a depth of 6 m. Agaricia agaricites is the dominant coral species, but Porites porites was dominant before Hurricane Allen in 1980. … two CARICOMP monitoring sites have been established in Barbados; one on the west coast just north of Holetown at Folkestone Park (13°11’18″N, 59°38’31″W), to monitor a nearshore hard coral reef (Bellairs fringing reef), and one on the southwest coast at Graeme Hall, St. Lawrence (13°04’06″N, 59°34’39″W), to monitor a mangrove wetland (Graeme Hall Swamp) and a seagrass bed (St. Lawrence Lagoon). …Graeme Hall Swamp covers an area of 32 ha in one of the most densely populated areas of the island (Riven-Ramsey, 1988) (Fig. 2). Residential development surrounds the swamp along the southern, eastern, western and northwestern boundaries, a main coastal road (Hwy 7) runs between the swamp and the sea on the south side, and agricultural lands border the swamp along the northeastern side. The swamp is essentially divided into an eastern and a western quadrant by a wide, man-made footpath/ roadway oriented from north to south through the area.  Fig. 2. Habitat map of Graeme Hall Swamp showing main vegetation types and location of CARICOMP monitoring plots. ..Graeme Hall Swamp drains directly into St. Lawrence Lagoon (Fig. 3). The lagoon covers an area of approximately 18 ha, is protected from high energy waves to seaward by an old reef flat and rubble bank (50-150 m wide), and becomes exposed at spring low tides. Extensive areas of sediment accumulation occur on the southwestern shelf of Barbados (Murray et al., 1977), and the lagoon is bounded on the landward side by an actively accreting, fine, coral sand beach (now 60 m wide). The lagoon is shallow (1-3 m deep at low tide); it has a sand substrate and seagrass beds that are occasionally exposed at spring low tides. The seagrasses are primarily turtle grass (Thalassia testudinum) and manatee grass (Syringodium filiforme), although some sparse, monospecific beds of shoal grass (Halodule beaudettei) occur nearshore in the vicinity of the mangrove swamp drainage canal (Oxenford et al., 1993). Macroalgae, Bryothamnion spp., Caulerpa spp. and Padina spp., are also present in the area directly adjacent to the swamp outfall. The seagrass beds act as adult foraging and/or nursery habitat for sand dollars (Mellita sexiesperforata), a commercially important sea urchin (Tripneustes ventricosus), the endangered green turtle (Chelonia mydas), and several estuarine and reef-associated fishes such as mullet (Mugil spp.), wrasse (Halichoeres spp.), razorfish (Xyrichtys spp.), and grunts (Haemulon spp.) (Oxenford et al., 1993). The CARICOMP monitoring sites are located in the main area of mixed seagrass towards the eastern end of the lagoon.
The St. Lawrence Lagoon is an important ecosystem to Barbados. It is the only area with significant seagrass cover on the west, southwest, and southeast coasts of the island. It also provides an important habitat for the commercially important sea urchin T. ventricosus, the critically endangered green turtle C. mydas, which feeds in the lagoon, and the hawksbill turtle Eretmochelys imbricata, which occasionally nests on the beach. It is also the only location in Barbados where mangrove swamps, seagrass beds, and deep hard coral reefs can be found in close association. A comparison of aerial photographs of the St. Lawrence Lagoon in 1964 and 1991 indicates that the area of seagrass cover has decreased and beach width has increased over the 27 years. In fact, in just the last few years the beach has accreted so rapidly that seagrass in the western end of the lagoon has been smothered by sand and replaced by a beach exposed at low tide. The lagoon is currently used as a mooring basin for small fishing boats and for recreational swimming and windsurfing. A proposal has been submitted to the Government to incorporate the area into a national marine park (Oxenford et al., 1993). |

| From Lotus Vermeer, 2000. Changes in seagrass growth and abundance of seagrasses in Barbados, West Indies, PhD thesis, Dalhousie University.
Ch 3: EFFECTS OF SHORT-TERM CHANGES NI COASTAL WATER QUALITY ON SEAGRASS ABUNDANCE AND LEAF GROWTH IN BARBADOS Ch 5: CHANGES IN ABUNDANCE AND LEAF GROWTH OF SEAGRASSES NI THE EASTERN CARIBBEAN: 1969-1994 |
| From Caribbean-Wide, Long-Term Study of Seagrass Beds Reveals Local Variations, Shifts in Community Structure and Occasional Collapse Brigitta A van Tussenbroek et al., 2014 in Plos 1
This study presents the results of a long-term (1993–2007, with some continuing to the present) Caribbean-wide seagrass monitoring initiative: the Caribbean Coastal Marine Productivity (CARICOMP) program….The CARICOMP program has generated a Caribbean-wide dataset using a simple, low-cost but standardized sampling protocol, consistent among sites over time…The seagrass communities at the majority (25) of the 35 stations included in the analysis for longer-term trends in the community showed changes in at least one of the six selected parameters (Table 1, Table S6). At six stations the seagrass beds collapsed: in Bermuda (3 stations) the decline was due to excessive grazing by sea turtles; in Barbados (2 stations) poor water quality followed by a population explosion of sea urchins and subsequent storms were responsible; and in Mexico, a coastal bed (1 station) was buried by sediments during a hurricane. Most monitoring stations (46 out of 52) were exposed at least once to a major meteorological event (hurricane or tropical storm, Table S1) during the study period, but apart from the above-mentioned collapse of communities in Mexico and Barbados (where the storms were not the main cause of collapse), minor impacts of storms were registered only at Belize-station 26 and Venezuela-stations 47 and 48 (Table 1). At 15 (43%) out of 35 studied stations (Table 1, Fig. 5), changes in the seagrass beds were consistent with hypothesized change scenarios of increased turbidity (Site 4-Stations 8 & 9 and Site 21-Station 49) or increased nutrient input (Site 2-Station 5; Site 5-Stations 10 thru13; Site 8-Station17; Site 10-Station 21; Site 12-Station 25; Site 14-Stations 33 & 34 Site 20-Stations 47 & 48). Most stations that showed shifts in community structure consistent with environmental degradation were reported to have received little or only moderate human-induced impacts at the onset of the study (Fig. 6). 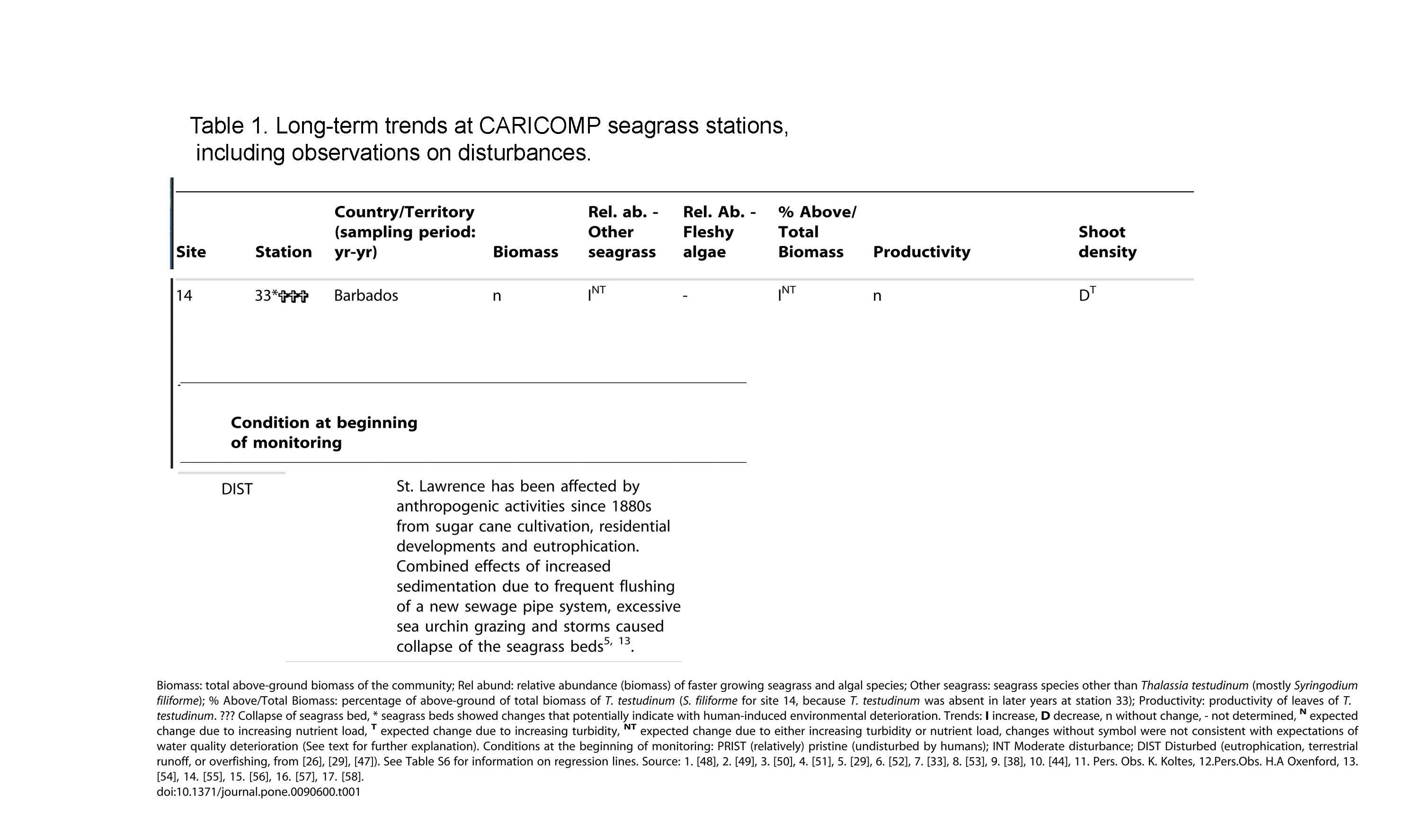 Screencapture extracts from Table 1. Click on image for larger version …The fate of the seagrass beds at Barbados-St. Lawrence lagoon (site 14) is a good example of how long-term (chronic) anthropogenic stress can act synergistically with acute extreme disturbance events [over-grazing by an exceptionally strong recruitment of sea urchins, Hurricane Ivan (2004) and Tropical Storm Emily (2005)] to cause collapse of an ecosystem. Even after 7 years, this lagoon has shown only minimal recovery, with just a few impoverished T. testudinum plants in areas of coral rubble and a very sparse vegetation of Halodule wrightii appearing in the sand areas (H. Oxenford, unpublished data). |