PRELIMINARY OBSERVATIONS OF THE STATUS OF SHALLOW WATER REEFS AT SANDY ISLAND, CARRIACOU, GRENADAA Report to the Grenada Board of Tourism & the Kido Project Environmental
Station
|
  |
MAJOR FEATURES
The elkhorn coral framework
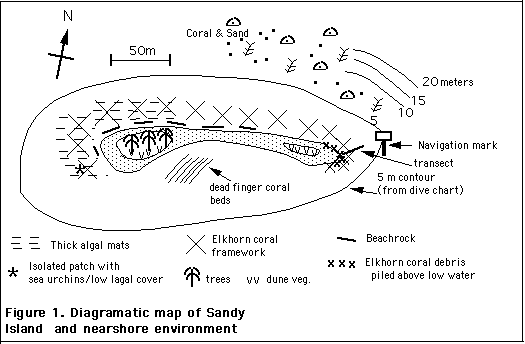
The most prominent feature of the shallow waters (0-4m) surrounding the island, is the occurrence of an "elkhorn coral framework " in a distinct band or zone on the east, north and west flanks of the island (Fig. 1, 2). This framework consists of completely dead, structural remains of elkhorn coral, Acropora palmata (photo plate II). The zone extends normal to shore over a distance of approximately 20-50 m, at depths of approximately 0.5 to 3.5 m (depth to the top of the framework). The elkhorn coral framework is separated from the beach by a shallow, sandy or sand/loose rubble bottom of a few to approximately 10m width except around the eastern end of the island; there, consolidated (bound) elkhorn coral rubble has accumulated above the low water line forming an emergent, rampart-like structure.
This distribution corresponds approximately to the reef crest zone of healthy reefs. Elkhorn coral was commonly the only significant species in the reef crest zone through the Caribbean prior to the Caribbean wide die-off of elkhorn coral in the late 70's/early 80's associated with an epidemic of "White Band Disease" (see Background, above). In 1969 living elkhorn coral formed characteristic dense, thickets of interlaced branches in the most exposed areas at Sandy Island (photos II C, E); in 1996, such thickets were still largely intact in some areas (photo IIF) , while in others, they had been largely disrupted (photo IID).
Except on the east/northeast flank of the island (described below), there are no living elkhorn corals in the elkhorn coral framework, and few other living or dead corals, except for some fire coral (Millepora spp.) In the northwest flank, there are some large, healthy colonies of lobed boulder coral, Montastrea annualaris (Plate III H) bordering seaward oriented channels which form breaks in the continuity of the elkhorn coral framework. These channels are regions of strong seaward return flow of water during storms.
PLATES
 |
PLATE I
|
 |
PLATE II
|
 |
PLATE III
|
New elkhorn coral and associated biotic gradients in the elkhorn framework zone.
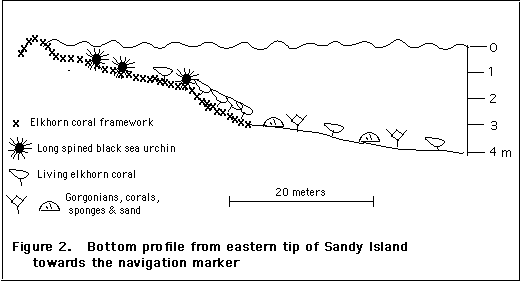
Over most of the elkhorn framework zone in 1996, there are no signs of living elkhorn coral. However, living colonies were found on the eastern flank of the island (Fig. 2; photos III D, E). The largest individual colonies are 75 cm in maximum surface dimension and there is a full range of smaller colonies. (This compares to maximum surface dimensions up to several meters in 1969). These colonies appeared completely healthy, and as yet, do not exhibit, old, encrusted growth, thus they appear to be of recent origin. Actively growing elkhorn coral occurs in greatest abundance and most regularly on the slope going from 1.5 to 3 m (Fig. 2). Occasional individual colonies or localized aggregations of living colonies occurred on the old elkhorn coral framework shoreward of the slope, where few other corals except for fire coral are found, and they also found mixed with other coral species on the coral-sand bottom seaward of the slope (Figure 2). In the areas of greatest density, intermingling of branches from different colonies forms an almost complete canopy. Numbers of elkhorn corals in five successive 2 x 2 m quadrats placed along the contours of the slope were 6,1,1,3,4,4.
Living elkhorn corals up to 1 meter maximum linear dimensions were also observed adjacent to Carriacou island proper at Anse La Roche (photo II F), which is described in a separate report (17). Other locations were not examined.
In the same area that the living elkhorn coral was observed on the southern flank of Sandy Island, long spined black sea urchins (Diadema antillarum ) are also abundant, and algal growth, which is dense elsewhere, is sparse. The black sea urchin is an important grazer of the algae that grow on old, dead portions of coral.; it was killed off by a Caribbean wide epidemic in 1983/4, and has only recently begun to return in high numbers and in only some areas (see Background, above) . Numbers of sea urchins observed in six 2 x 2 m quadrats were 10, 6, 2, 5, 3, 7, 8. (These numbers refer to sea urchins readily observed; they are low estimates of total numbers as more urchins were readily found on more careful examination of the cavitous framework).
The density of the living elkhorn coral decreases with increasing distance west along the north side of the Sandy Island, reaching near zero at about one-third of the distance towards the west end of the island. Frequency of patches of black sea urchin also decrease in frequency in the same direction. Algal cover, in contrast, increases, from very little on the dead elkhorn coral on the east flank (photo III D, E) to a uniform "fuzz" at one third of the distance towards the western extremity of the island, and to heavy overgrowth at the western extremity of the elkhorn coral framework (photos III A, B).
In the area of heaviest overgrowth of the old elkhorn coral by algae (Fig. 1), algal cover is 100% and the algae form a dense turf up to several centimeters thick (photo IIIB) which in some areas is topped by bushy growth usually characteristic of high nutrient systems. There are 5+ species common in this assemblage including Digenia simplex, Ulva (close to shore), an unidentified filamentous alga, Laurencia obtusata, and delicately branched Amphiroa sp. and Centroceros sp. (Please note that this description is based on field examination of only a few samples).
There is a notable absence of the sand-forming Halimeda species through the entire area. Correspondingly, Halimeda platelets are not a common constituent of the sand on the island.
Over most the elkhorn coral framework at Sandy Island, the old, dead elkhorn coral fragments making up the framework, while mostly no longer in growth position, are firmly bound together by encrusting coralline algae. Where this is the case, further disruption of the existing framework will probably be slow. However, in the northwestern and western flanks, the existing framework - some if it consisting of dead colonies that are still in growth position and some of redistributed material - could be pulled apart by hand.
Thus there appears to be an overall positive correlation between occurrence of living elkhorn coral, sea urchins, binding by coralline algae, and paucity of mat and fleshy algae.
Close to southwest extremity of the elkhorn coral framework, where algal cover is very heavy, a single patch about 5 m diameter was found that was largely clear of algae. Black sea urchins were observed in the patch, but not outside of it, thus, the sparsity of algae in the patch is attributed to grazing activity of the black sea urchins. The coral debris within this patch was not in growth position, and appeared to have been physically disturbed (possibly thrown around by wave action or disturbed by traps which are set in this area) at some time.
Surgeonfish and parrotfish are important herbivorous fish species in the Caribbean (18). While at the site, several large schools of surgeon fish were observed on the eastern flank. Small to medium sized parrotfish were common towards the southern border of the old elkhorn coral framework on the west flank of the island, and both surgeon fish and parrot fish were observed in lower numbers elsewhere.
Communities beyond the elkhorn coral framework
Beyond the elkhorn coral framework, in depths greater than 3-4 m, a coral-sand bottom slopes gently to deeper water; it includes many gorgonians (sea whips, sea fans), brain corals, starlet coral (Porites asteroides ), occasional elkhorn coral, boulder coral (Montastrea annularis) and the finger coral (Porites porites ), giving 5-30% coral cover (photo III G). There are also interspersed beds of finger coral which give 50-60% cover in some regions. Overall, these coral-sand communities looked healthy, and were without notable diseases on living corals, or significant overgrowth by algae. Occasional, completely dead but intact brain corals were observed; these may have been killed by black band disease.
The anchorage in the lee of the island
The sea bottom on the south, protected side of the cay where yachts anchor is sand and rock, with some completely dead beds of finger corals. There is a heavy cover of algae over most of the bottom. These include a bushy alga (Acanthophora spicifera.) and soft filamentous algae; towards the west this cover blends into the algal cover on the elkhorn coral framework. Overall, this bottom is unattractive. The heavy algal growth is suggestive of localized nutrient and organic pollution.
Currents
While at the site, there was a strong current flowing from east to west beginning at about 10 m from shore to the north and south of the island, and impacting on shore on the eastern flank. The current was strongest in the area of the highest density of living elkhorn corral and high sea urchin density, on the east flank. The west flank was mostly in the lee of the island in regard to current; there was little water motion in that area and water was noticeably warmer than elsewhere.
According to Carriacou dive operator, Tankred Mueller, the current at Sandy island normally flows NE to SE, and rarely flows in the reverse direction, perhaps for only 5 days a year. Swells normally approach from the north or northeast. He comments that when swells come from the north or northwest, or worse, from the west, there is direct damage. Reportedly in October 1992, the northwest swells from a storm hit Sandy for some days, eroding drastically the base and bringing down the remaining 8 grown palm trees on the "eastern cay", and removing a considerable chunk of beach off the northwest side of the "western cay". The sand was slowly brought back by wave action. He comments reports that at depths 7-8 meters on the northwest of Sandy there was up to 40 cm less sand (he measured it because he has a permanent dive mooring buoy and inspected its foundations). Tankred reports that NW ground seas can be felt at depths of 20 m.
CONCLUSIONS
Below, the observations above are interpreted with reference to the scientific literature to arrive at five main conclusions (highlighted).
(i) The die-off of elkhorn coral, and the absence to date of substantial renewal or replacement of this species are attributable to factors beyond local control (mass moralities of elkhorn coral, and subsequently of black sea urchins, in the late 70's/early 80's); however, local factors (nutrient/organic pollution from yachts and overfishing) may have exacerbated the problem.
The complete degradation of the elkhorn coral zone that was flourishing and healthy in 1969 (photo plate II) is attributable to two major events that were not of local origin. First, there was the mass mortality of elkhorn coral associated with White Band Disease which ravaged elkhorn coral throughout the Caribbean over a roughly 8 year period the late 70s/early 80's (19). The scale of the die-off at many locales (>99%) eliminated the usual source of "seed" for new colonies when exceptional storm events cause great damage viz., broken fragments of elkhorn coral. Regeneration of the community was then dependent on larval settlement and subsequent growth which is much slower than regeneration from fragments.
Second, following closely on the heels of the first event, there was mass mortality of black sea urchins, associated with a disease which swept the Caribbean over an even shorter interval (roughly one year) and caused greater than 93% mortality throughout the Caribbean (20). Research conducted prior to this event had suggested that the black sea urchin plays a keystone role in coral reef communities in the Caribbean through its continual grazing down of algae growing on old coral. Coral and algae compete for space, nutrients and light; the algae are fast growing and capable of quickly outgrowing corals, however in coral-dominated communities, such growth is restricted by low nutrient supply (corals are better able to grow under low nutrient conditions) and by grazing of herbivorous invertebrates and fish.
When sea urchins were removed experimentally from areas where herbivorous fish stocks were low due to overfishing, algal growth increased many fold, encroaching on coral colonies and reducing settlement of coral larvae. Predictably, mass moralities of sea urchins in 1983/4 was followed by sharp increase in algal cover, except in locales where there were still large populations of herbivorous fishes to compensate for loss of urchins (15).
It appears that once established, an algal dominated system may be quite stable, as a dense cover of algae reduces settlement of coral larvae, and even settlement of sea urchin larvae (21) ; further there may be selection of species or forms of algae that are structurally or chemically resistant to grazing by the fishes (which otherwise would not be limited by settlement and slow movement as are sea urchins) (22). The result is that the change in state initially brought about by catastrophic disturbances of short duration, can persist well beyond the causative disturbance(s)(23), creating "an alternative stable state"(24).
While the major factors leading to deterioration of the elkhorn coral at Sandy Island were not of local origin, overfishing and nutrient pollution which are under local control, may have exacerbated the problem. Stocks of parrot fish and surgeonfish at Sandy Island appear to be well below those that would occur on an unfished or moderately fished reef, presumably due to intensive fishing, but selection of resistant forms of algae may also be involved.
Algal growth of the intensity that occurs on the elkhorn coral in the west flank is usually associated with nutrient enrichment whether natural or artificial (25). These nutrients could emanate directly from yachts, or from polluted sediment in the region where yachts anchor; the heavy algal growth in that area is suggestive of nutrient pollution. Nutrients may be moved into the western flank area by eddies breaking off from the westward moving currents; released into the quiet waters, they would reside there long enough to be taken up by algae, stimulating growth. A very graphic example of nutrient stimulated algal growth and associated coral death can be observed at nearby Jack Adam Island; there, conch shells have been discarded in recent years forming large mounds from which there is seepage of putrid material (photo IE) into the adjacent submarine where rocks are covered by heavy growth of algae and corals have died.
(ii) Increased erosion of the island in recent years is attributable to the degradation of elkhorn coral beginning sometime in the late 70's/early 80's; in the absence of regeneration of elkhorn coral, most of what remains of Sandy Island will probably be lost
Living elkhorn coral thickets such as those shown in Photos II C & E greatly reduce wave energy in the lee of the thickets. Normally these thickets are dynamic structures, continually being added to by new growth, and detracted from through continuous activity of boring organisms and physical fragmentation. Exceptional storm events can cause extremely large losses of living elkhorn coral (26) however, in many or most situations they have reestablished in the longer term, as evidenced by the occurrence of elkhorn coral as a major constituent in limestones (27). Indeed, Sandy Island likely owes its existence to the persistence of elkhorn coral for many hundreds or thousands of years.
In a healthy reef, establishment of new colonies of elkhorn coral, for example after extensive storm damage, occurs mainly by growth of fragments broken off from preexisting colonies (28). The White Band Disease effected complete or very nearly complete mortality of elkhorn coral, eliminating this source of new coral. Regeneration from larval settlement has been very limited so far and other species have not moved in to fill the niche once occupied by elkhorn coral. Thus for 12+ years, there has been virtually no additions to balance losses.
Growth of encrusting coralline algae appears to have stabilized the old framework on the east flank and over much of the north flank of the island. Thus the island area to the lee of the east and northeast flanks, may at this point be relatively stable. However towards the west flank, and on the west flank, the heavy growth of mat-forming and bushy algae has precluded growth of encrusting corallines, and the framework there is not firmly bound together. Hence it could be expected that the framework in that area - which still retains a lot of the original structure - will be progressively punctured and that the adjacent land mass will still be subject to heavy erosion when strong wave approach from the west or northwest. Residents report that recent losses have been most severe on the northwest part of the island, which is consistent with this concept.
In the absence of regeneration of elkhorn coral, it seems a pretty safe prediction that most of what remains of Sandy Island will be lost, that this loss will occur mostly over short intervals during exceptional storms, rather than gradually, and that the losses will become increasingly large relative to what's left with each event. In this context, Sandy Island is in a very precarious condition.
(iii) There are encouraging signs of recovery of elkhorn coral at Sandy Island; this recovery is linked to recolonization and maintenance of low algal cover by black sea urchins
The size distribution and morphology of the living elkhorn corals on the eastern flank are similar to those described by Jordan-Dahlgren on the Veracruz reef complex in a 1993 paper on "Recolonization patterns of Acropora palmata in a marginal environment"(29). He suggested that the largest colonies at that site, which were about 1 m in maximum linear dimension, could date back to a mortality event 20 years previous, assuming a growth rates of 8-9 cm per year, and thus could be survivors of the old colonies, or have arisen from larval settlement. The more numerous, smaller colonies, he suggests, were only a few years old, and must have arisen from larval settlement. The maximum colony dimension at Sandy Island was about 75 cm, but the maximum linear growth dimension was less than that, probably about 40 cm. Thus these colonies are possibly a maxmimum of only 5 years old, and are mostly less than 5 years. This population has likely developed relatively recently through settlement of larvae carried to the island by currents.
Larval settlement in Acropora species is strongly suppressed by algae (30) . The consistent co-occurrence of black sea urchins in areas where living elkhorn corals were observed, and of sea urchins in all areas where algal cover is low (with or without elkhorn coral) suggest that recolonization of old coral by sea urchins is a prerequisite for larval settlement and subsequent growth of elkhorn coral. Photographs taken in 1969 also illustrate the occurrence of black sea urchins in elkhorn coral stands, and little algal growth on old coral (Photo IIA).
There appears to be some relationship of re-establishment of sea urchins to currents; the area of elkhorn coral and high sea urchin density occurs on the upstream, eastern flank of the island where there is a consistent current onto the slope. The one patch of sea urchins found on the west flank, lay on the edge of the westward moving current south of the island. It can be hypothesized that areas impacted directly by currents have greater influx of larvae. On the east/northeast flank, where algal cover is low over a broad area, it is possible also (or as an alternative hypothesis) that nutrient pollution was minimal in this upstream area compared to other areas around the island, even before sea urchins began to return, resulting in greater survival of settled larvae than in areas with heavier algal cover. The fact that encrusting coralline algae are common through the east/northeast flank may indicate that this area has always had less algal cover than on the northwest/western flank.
(iv) There has been extensive anchor damage of coral communities at Sandy Island
Whole beds of finger coral on the leeward side of the island where yachts anchor were observed to have been completely disrupted by anchors. Partial damage of finger coral beds was noted in other areas which is probably due to anchors form smaller boats. Anchor damage and boat groundings associated with tourist traffic are cited as a major cause of reef decline in the Caribbean (31); Sandy Island is no exception.
(v) The coral communities beyond the elkhorn coral framework to the east and north of the island, at depths of 3-4 m and greater, appeared to be in generally good condition, however, more critical examination is needed to properly asses their health.
In general , there appears to have been less degradation of deeper water reefs than of shallower reefs in the Caribbean (32).
ACCELERATING THE RECOLONZATION BY ELKHORN CORAL
As discussed above, the algal-dominated state, once established, tends to resist reversion to the original state because the heavy algal growth prevents settlement of invertebrate larvae, including those of sea urchins; further and there may be selection for forms and species of algae that not edible by the more mobile fish grazers. Coral debris in the one patch of sea urchins/low alga cover observed on the west flank appeared to have been physically disturbed at some time; this could have exposed fresh surface not covered by algae, which would favor settlement and survival of sea urchin larvae. The patch had sharp boundaries with uniformly high algal cover outside of it, and low algal cover within it; this suggests that once established within a high algal region, sea urchins are capable of maintaining low algal cover within the patch and presumably, of expanding the patch at its edges as the population grows.
We propose that the recovery of elkhorn coral at Sandy Island could be accelerated by establishing a number of such "recovery patches" in the areas presently lacking black sea urchins and living elkhorn coral. In each patch, algae would be scraped off of the old coral debris, and black sea urchins introduced simultaneously to the patch; when the black sea urchins have proven to be established and to limit algal regrowth, pieces of living elkhorn coral would be transplanted into the site and stabilized in their new positions by use of cement or epoxy.
Efforts to reestablish coral communities by transplanting have been successful in some areas or with some species, but not in others (33). Two factors are in favor of it being successful with elkhorn coral at Sandy Island:
(i). Elkhorn coral normally exhibits vigorous asexual regeneration (regeneration by regrowth of fragments broken off from existing colonies) (34), and , the species has been found to very amenable to transplanting (Dr. D. Hubbard, USVI, personal communication).
(ii). When numbers of any species get very low, there is a high probability of going extinct (or locally extinct) because catastrophic events can completely wipe out the few individuals remaining. New recruitment of elkhorn coral has been observed at other sites in the Caribbean, but they have been repeatedly set back by hurricane damage (Dr. W. B. Gladfelter, USVI, personal communication). The Grenadines lie outside of the main hurricane belt, which greatly increases the odds of an experiment of this sort being successful.
There are some uncertainties about whether newly established elkhorn coral populations, whether established by natural or artificial means, will be sustained. The new colonies at Sandy Island were likely established by settlement of larvae produced elsewhere. We do not know that source. Studies have indicated that sexual reproduction by elkhorn coral is seasonal (35). Regardless, new colonies of the coral are now present, and can be replicated by asexual reproduction, i.e. taking fragments and transplanting them. New colonies were observed also at Anse La Roche in Carriacou, suggesting that for the moment there is a significant influx of larvae of this species coming into the nearshore waters of Carriacou.
Another uncertainty concerns the possibility that new populations will again be wiped out by White Band Disease. The disease has been seen to reoccur since the epidemic die-off (36) but it is possible that some or all of the new recruits would be resistant to it, and/or that conditions that were conducive to its epidemic development in the 1970's and 80's will not reoccur. For the moment, the answer to the questions of whether white band could again strike and wipe out the population is simply unknown. Similarly the disease that caused the epidemic die-off of the black sea urchin could reoccur.
Regardless Dr. W. B. Gladfelter commented that it would be worthwhile to attempt to assist recovery, as we propose, especially given the more favorable odds with respect to hurricanes in comparison to other areas.
Increasing the odds
It is reasonable to expect the elkhorn coral to continue to recover and reestablish its canopy structure, however, given the still very low level of the population, the return could be seriously set back by chance events such as exceptional storms, or by human activity. Elkhorn coral is an environmentally sensitive species, requiring clean, clear water, and not very tolerant of low salinity or suspended sediment. The species has been lost in regions affected by human habitation, well before the modern decline (37); increased sediment load is often the major factor as this species lacks effective sediment removing mechanisms. Establishing accelerated recovery centres is one way of significantly increasing the odds in favor of regeneration of the elkhorn community at Sandy Island; another, is to minimize existing or potential negative human impacts on the system, including
(i) physical damage due to anchors and boat groundings;
(ii) physical damage due to snorkellers and divers: if the recovery of elkhorn coral at sandy Island becomes a matter of public interest, it will be important to encourage people not to approach the living colonies which are readily broken by stepping on them;
(iii) nutrient/organic pollution from yachts and shore parties;
(iv) severely reduced stocks of herbivorous fishes due to overfishing;
(v) sedimentation: any activities in the vicinity of Sandy Island that could increase the sediment load (or load of nutrient or toxic materials) in water flowing onto elkhorn coral should be avoided.
It seems worthwhile to attempt to reestablish the elkhorn coral at Sandy island because it may the only chance to do that. If successful, this could lead to the techniques being used elsewhere in the Grenadines, essentially reversing what otherwise would be a long term loss with very considerable ecological, geological and probably economic impact.
Accelerating the process of return of sea urchins and control of algae, even without return of elkhorn coral, could be of benefit by allowing more growth of encrusting coralline algae. That would strengthen the old framework, and may provide a base for growth of other coral species as appears to have occurred on the west coast of Barbados following decline of elkhorn coral associated with human settlement (38).
Regardless of the outcome, this exercise provides a focus for people to learn about and become sensitive to the ecology of the coral reefs and to learn techniques that might be used in conservation and restorative work with other species and at other locations.
RECOMMENDATIONS
1. Steps should be taken to reduce as much as possible, existing or potential negative impacts of high tourist traffic on the reefs at Sandy Island. These include primarily damage from boat anchors, boat groundings, foot damage from snorkellers and divers, and nutrient/organic pollution.
Installation of moorings as recommended by the Save Sandy Island Plan addresses the issue of anchor and boat grounding. Physical damage from deliberate or inadvertent handling of corals by snorkellers and divers is especially of concern in relation to the new elkhorn coral growth. Other of the Save Sandy Island Plan's recommendations provide some structure for dealing with this issue, viz. their proposed management initiatives 2,8,10. The issue of nutrient./organic pollution from yachts needs also to be specifically addressed. A combination of public education and regulation may be required to deal effectively with this issue.
2. Some regulation of fishing in order to increase stocks of herbivorous fishes should be considered.
There are a number of options that could be considered, e.g. a complete fishing ban, rotational harvests (fishing allowed in only certain sectors at any one time), fishing zones, regulation of mesh size, species restrictions. As current stocks are probably below "maximum sustainable yield", some regulation could actually increase the stocks and the sustainable harvest at the same time (39). Increased stocks would also increase the ecotourism value of the island. The fragility of the reef and the need to site traps and secure them properly to minimize physical damage to the reef is also a factor to be considered.
3. Care should be taken to avoid operations in adjacent waters that might pollute the water flowing onto the Sandy Island reefs with nutrients, toxic chemicals or increase its sediment load.
Elkhorn coral is particularly sensitive to water quality problems of this nature.
4. Formal scientific documentation of the Sandy Island site and trials of the proposed "accelerated reef recovery technique" would be valuable scientifically; by combining a scientific study with training of local personnel in coral reef ecology, and using the information and skills to make Sandy Island a recognized site for ecotourists, the tourism industry and fishers to learn about coral reef ecology, the "experience of Sandy island" could have benefits well beyond its boundaries.
The descriptions provided in this report, while a reasonable basis for some management initiatives, are only preliminary ones, scientifically. They are a basis for designing a proper scientific study. From that perspective, it would be appropriate to document quantitatively and further elaborate on the qualitative observations given in this report, set up permanent monitoring transects and quadrats extending to deeper water, and to conduct replicated experiments to test the inferred relationships.
The reefs at Sandy Island in themselves constitute a very small proportion of the total reef frontage in Carriacou, and are not particularly important for protection of the mainland (Carriacou proper), although other reefs at Carriacou are in this regard. Obviously, detailed scientific studies cannot be conducted for every inch of coral reef that needs to be managed. The value of doing this at Sandy Island lies in the more general value that would have as a scientific model, and coupled with community based activities, as an educational tool.
Coral reef researchers have emphasized that the support and involvement of communities is required for sustainable coral reef management (40). Sandy Island has already focused attention of islanders on the issue of coral reef health, it is readily accessible, presents the major types of problems of exotic and local origin that have caused degradation of Caribbean reefs, and offers an opportunity for testing/developing some novel approaches to coral reef restoration. Developed in this context, restoration of the Sandy Island reefs could stimulate restoration and conservation of coral reefs more broadly in the eastern Caribbean, and provide new economic opportunities for Carriacouans.
"The conservation of coral reefs and other shallow tropical marine ecosystems depends upon the outcome of a race between the accelerating degradation of these ecosystems and two related areas of human endeavor: the development of ecologically and sociologically sound models for management and the effective education of people to the value of biological conservation" (40).
FOOTNOTES
- "Carriacou's Sandy Island. An Asset to Tourism and a Liability to the Environment", in December 1995 Issue of the Caribbean Compass, p. 12.
- Prof. Patriquin conducted some of his PhD research on seagrass communities in Carriacou in 1968 and 1969, and in 1994 resurveyed the same sites in collaboration with Dr. Hunte as part of a program for monitoring long term changes in coral reef systems. Prof. Patriquin had also visited Sandy Island in 1969 and took underwater photographs of the then thriving Acropora palmata dominated reefs, included in this report. Dr. Hunte has researched long term changes in reefs at Barbados and elsewhere in the Caribbean with a particular interest in how the interactions between herbivores, algae and corals influence variation in coral cover over space and time.
- Where not specifically identified, supporting documentation for this section can be found in (i) Ginsburg, R.N. (Complier). 1994 Proceedings of the Colloquium on Global Aspects of Coral Reefs: Healtg, Hazards and History, 1993, Rosentiel School of Marine and Atmosphereic Sciences, University of Miami. (ii) Sorokin, Y.I. 1993 Coral reef Ecology. Springer-Verlag, Berlin, New York. (iii) Caribbean Conservation Association, 1991. Country Environmental Profile: Grenada. Caribbean Conservation Association, Barbados.
- Ward, P. 1995. The End of Evolution. Bantam Books.
- Kojis, B.L. & Quinn, N.J. 1994 Biological limits to Caribbean reef recovery: a comparison with western south Pacific reefs, pp 273-278 In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 353-359.
- (i) Scoffin, T.P. 1994. History of a fringing reef on the west coast of Barbados 1974-1992. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 273-278 (ii) Woodley, J.D. et al., 1981. Hurricane Allen's impact on Jamaican coral reefs. Science 214: 749-755.
- Goreau, T.J. & Hayes, R.L. 1994. Coral bleaching and ocean "hot spots". Ambio 23: 176-180.
- Kojis, B.L. & Quinn, N.J. 1994 Biological limits to Caribben reef recovery: a comparison with western south Pacific reefs. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 353-359.
- Hubbard, D.K., Gladfelter, E.H. & Bythell, J.C. 1994. Comparison of biological and geological perspectives of coral reef community structures at Buck Island, U.S. Virgin Islands. In : Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 201-207.
- Sheppard, C. 1993. Coral reef environmental science: dichotomies, not the Cassandras, are false. Reef Encounter 14 (October): 12-13.
- (i) Richardson, L.L. 1996. Horizontal and vertical migration patterns of Phormidium corallyticum and Beggiatoa spp. associated with Black-Band Disease of corals. Microbial Ecology 32:323-335. (ii) Edmunds, P.J. 1991. Extent and effect of Black Band Disease on a Caribbean reef. Coral Reefs 10:161-165.
- Smith, S.R. & Ogden, J.C. (Editors). 1994 Status and recent history of coral reefs at the CARICOMP network of Caribbean marine labortaories. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 73-78.
- (i) Allard, P. 1993. Changes in Coral Community Structure in Barbados: Effects of Eutrophication and Reduced Grazing Pressure. MSc thesis in Biology, McGill University, Montreal. (ii) Smith, S.R. & Ogden, J.C. (Editors). 1994 Status and recent history of coral reefs at the CARICOMP network of Caribbean marine laboratories. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 73-78.
- Coral reef degradation: a long term study of human impacts. In: Ginsburg, R.N. (Compiler) 1994 cited in (3) above, pp 208-212.
- Lessios, H.A. 1988. Mass mortality of Diadema antillarum in the Caribbean. Annual Reviews of Ecology and Systematics 19: 371-93.
- Caribbean Conservation Association, 1991. Country Environmental Profile: Grenada. Caribbean Conservation Association, Barbados; Smith, S.R. & Ogden, J.C. (Editors). 1994 Status and recent history of coral reefs at the CARICOMP network of Caribbean marine laboratories. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 73-78.
- Patriquin, D.G. 1996. Preliminary observations of Anse La Roche beach June 20, 1996, and comments on the status of the beach as a site for ecotourism. Mimeogr. report to Grenada Board of Tourism and Kido Project Environmental Station.
- Smith, S.R. & Ogden, J.C. (Editors). 1994 Status and recent history of coral reefs at the CARICOMP network of Caribbean marine laboratories. In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 73-78.
- Gladfelter, W.B. 1982. White Band Diseases in Acropora palmata: implications for the structure and growth of shallow reefs. Bulletin of Marine Science 32: 639-643.
- Lessios, H.A. 1988. Mass mortality of Diadema antillarum in the Caribbean. Annual Reviews of Ecology and Systematics 19: 371-93.
- (i) Wittenberg, M. and Hunte, W. Effects of eutrophication and sedimentation on juvenile corals. 1, Abundance, mortality and community structure. Marine Biology 112:131-138; (ii) Hunte, W. and Wittenberg, M. 1992. Effects of eutrophication and sedimentation on juvenile corals. II. Settlement, Marine Biology 114:625-631.
- Duffy, J.E. & Hay, M.E. 1990 Seaweed adaptations to herbivory. BioScience 40: 368-375.
- Steneck, R.S. 1994. Is herbivore loss more damagung to reefs than hurricanes? Case studies from two Caribbean reef systems (1978-1988) In: Ginsburg, R.N. (Complier) 1994 cited in (3) above, pp 220-226.
- Hatcher, B.G. 1984. A maritime accident provides evidence for alternate stable states in benthic communities on coral reefs. Coral Reefs 3: 199-204.
- Littler, M.M., Littler, D.S. & Titlyanov, E.A. 1991. Comparisons of N- and P-limited productivity between high granitic islands versus low carbonate atolls in the Seychelles Archipelago: a test of the relative dominance paradigm. Coral Reefs 10: 199-209.
- Woodley, J.D. et al., 1981. Hurricane Allen's impact on Jamaican coral reefs. Science 214 :749-755.
- (i) Lewis, J.B. 1984. The Acropora inheritance: a reinterpretation of the development of fringing reefs in Barbados, West Indies. Coral Reefs 3: 117-122. (ii) Jackson, J.B.C, 1992. Pleistocene perspectives on coral reef community structure, American Zoologist 32:719-731.
- Highsmith, R.C. 1982 Reproduction by fragmentation in corals. Marine Ecology - Progress Series 7: 207-226.
- Jordan-Dahlgren, E. 1992. Recolonization patterns of Acropora palmata in a marginal environment. Bulletin of Marine Science 51: 104-117.
- Tanner, J.E. 1995. Competition between scleractinian corals and macroalgae: an experimental investigation of coral growth, survival and reproduction. Journal of Experimental Marine Biology and Ecology 190: 151-168.
- Allen, W.H. 1992. Increased dangers to Caribbean marine ecosystems: cruise ship anchors and intesnified tourism threaten reefs. BioScience 42: 330-335.
- Bak, R.P.M. and Nieuwland, G. 1995. Long-term change in coral communities along depth gradients over leeward reefs in the Netherlands Antilles. Bulletin of Marine Science 56: 609-619.
- Hatcher, B.G., Johannes, R.E. & Robertson, A.I. 1989. Review of research relevant to the conservation of shallow tropical marine ecosystems. Oceanography and Marine Biology Annual Review 27: 337-414.
- Highsmith, R.C. 1982 Reproduction by fragmentation in corals. Marine Ecology - Progress Series 7: 207-226.
- Szmant, A.M. 1986. Reproductive ecology of Caribbean reef corals. Coral Reefs 5: 43-54.
- (i) Dr. W. B. Gladfelter, USVI, personal communication. (ii) Bythell, J. & Sheppard, C.R.C. 1993. Mass mortality of Caribbean shallow corals. Marine Pollution Bulletin 26: 296-297.
- (i) Jaap, W.C. and Sargent, F.J. 1994. The status of the remnant population of Acropora palmata ( Lamarck, 1816) sat Dry Tortugas National Park, Florida, with a discussion of possible causes of changes since 1881. In: Ginsburg, R.N. (compiler), cited in (3), pp101-105. (ii) Lewis, J.B. 1984. The Acropora inheritance: a reinterpretation of the development of fringing reefs in Barbados, West Indies. Coral Reefs 3: 117-122.
- Lewis, J.B. 1984, cited in previous footnote.
- Russ, G.R. & Alcala, A.C. 1996. Do marine reserves export adult fish biomass? Evidence from Apo Island, central Philippines. Marine Ecology Progress Series 132: 1-9.
- Hatcher, B.G., Johannes, R.E. & Robertson, A.I. 1989. Review of research relevant to the conservation of shallow tropical marine ecosystems. Oceanography and Marine Biology Annual Review 27: 337-414.